Me to a friend: “I’m exhausted. I could really use a break.”
My friend’s response: “Which leg do you want me to break—your right or your left?”
Some might think that our exchange is funny, but it’s definitely equivocation. It’s equivocating over the meaning of the word “break.”
If you know me, you know I’d chuckle at my friend’s response and probably say, “Both of them—that way I can get some rest!” But most of the time I’m not too jovial about folks who equivocate over word usage when the topic is serious or related to science and discovery and unpacking life’s truths.
Words Matter
I believe we have a lot to learn from one another. I believe we get closer to the truth the more we listen to others and examine things from multiple angles. But if we aren’t using words to mean the same thing, communication will be frustrating at best. And if we never pause to clarify and unequivocate, we can never really communicate.
I care so much about communicating and not equivocating that I’m working on a book project related to unequivocating the word “evolution” for nonscientists. The book project takes me away from blogging. I’ve also been traveling a lot, and traveling makes blogging more challenging as well. There was NCCA in North Carolina, where I spoke on the topics of scientific data, interpretation, and storytelling and on unequivocating evolution. This was followed by a conference in Monterey, California, where I spoke on integrating science and faith and unequivocating evolution. And just two weeks ago, I was in San Antonio, Texas, for the annual Evangelical Theological Society meeting, where I spoke on having a Christian theology of viruses at a Chinese church for a joint RTB San Antonio chapter meeting.
I was reminded again at NCCA about how equivocation really exacerbates poor communication and leads to high levels of tension among clashing perspectives. This time the offending word was “novelty”—evolutionary novelty—in the context of a long-term evolution experiment.
Evolving Microbes
Maybe you’ve never heard of the long-term evolution experiments (LTEE) in E. coli started by a research group at Michigan State University in the laboratory of Richard Lenski. My colleague Fazale Rana has written about the experiments a few times over the past years, so maybe you have! (See “Resources” below.) The experiments began in 1988 when a single (clonal) bacterial culture was split into 12 separate cultures. The growth conditions have been carefully controlled, and samples are collected every 500 generations (every 75 days) to follow the evolutionary course of E. coli over time. Each culture has been continually grown in a glucose-limited medium in the presence of citrate and propagated via serial passage on a daily basis over the past 28 years. (Thank God this wasn’t a single graduate student’s project!)
One culture is of particular interest in this long-term evolution experiment because it has evolved a novel trait. By 31,500 generations, one culture overcame a growth restriction and began to (weakly) utilize citrate as a primary source of carbon and energy in the presence of oxygen.1 Prior to this, E. coli have been classically characterized by an inability to use environmental citrate under aerobic conditions.
The inhibition occurs because, in the presence of oxygen, a regulatory switch (a repressor) shuts down expression of the citrate transporter gene (citT) in E. coli. Under anoxic conditions, the citTgene is expressed, and E. coli produces a CitT protein that functions as a citrate/succinate antiporter, allowing uptake of citrate from the environment. Once internalized, citrate can serve as an energy source via a citrate fermentation cycle.2
However, under aerobic conditions, Cit+ E. coli (Ara-3) is able to utilize citrate from the environment due to the novel expression of a citrate/succinate antiporter (CitT). This acquisition of a new trait allows Cit+ E. coli to expand into a citrate-containing ecological niche that was previously not utilized. As a result, subsequent cultures (over 34,000 generations) in this lineage of the LTEE demonstrate greater biodiversity of E. coli than the ancestral cultures because they now contain two notable types of E. coli: those that utilize citrate as their primary energy source (Cit+) and a smaller subpopulation of E. coli that utilizes glucose at a much faster rate than the Cit+ bacteria. The authors rightly conclude that the LTEE allowed evolution of a novel trait in E. coli and led to an increase in biodiversity.3 These are hallmarks of evolution. And as such, the emergence of Cit+ E.coli is an excellent example of one type of microbial evolution.
Bacteria Accrue Genetic Mutations over Multiple Replication Cycles
During replication of the bacterial genome, replication errors may result in deletions, insertions, inversions, and various point mutations. Recombination and transposition of genomic segments also occur in bacteria and contribute to genetic variations. Although the processes can be described, the specific molecular triggers of these various genetic modifications are not always well characterized. They may be just a by-product of error-prone polymerases during high-speed replication (or transcription) in busy, tangled, and crowded intracellular environments. Such errors are often exacerbated under dystopian conditions such as starvation, heat shock, DNA damage, or other stress-inducing stimuli.
Mutations or large genomic restructuring that is detrimental will be quickly lost from the bacterial population, as bacteria with such altered genomes will not survive. Mutations that allow bacteria to continue to compete for energy/food sources will accrue and vary from bug to bug over time, without noticeable changes in the bacteria culture. Some unnoticeable mutations may potentiate other changes over time, and since they have no corresponding phenotype, they will be nearly impossible to identify. Although some early mutations may potentiate the bacterial genome for subsequent trait-altering mutations, they do not necessitate subsequent mutations. In other words, stepwise changes may eventually contribute to a final, distinct phenotype in a contingent fashion, but not all stepwise changes will lead to the same outcome or a change in phenotype.
Researchers in the LTEE have discovered that the Cit+ bacteria accrued such unidentified, potentiating mutations prior to generation 31,000, so by generation 31,500, a genome segment duplication of a portion of the citT operon occurred, actualizing the Cit+ phenotype. Additional mutations, including amplification of the duplicated segment,occurred in the Cit+ bacteria by generation 33,000, refining or fine-tuning its growth on citrate. All of these changes contributed to a new Cit+ lineage of E. coli and occurred in a stepwise and contingent fashion, not as rare simultaneous events of multiple or single mutations.4 This stepwise and contingent accumulation of contributing mutations is also a hallmark of evolution.
Are the Claims of Evolutionary Novelty Justified?
The 2012 report on Cit+ bacteria opens with this statement: “Evolutionary novelties are qualitatively new traits that open up ecological opportunities and thereby promote diversification.” According to the authors, the Cit+ E. coli underwent a “long-delayed and unique evolution” encompassing a “key innovation” involving multiple steps that resulted in a “new Cit+ function.” The research indeed shows that multiple, stepwise mutations were needed to potentiate, acquire, and refine the bacteria for citrate metabolism under aerobic conditions and that this one lineage of E. coli acquired this novel function in a contingent fashion.
Importantly, the authors claim that the evolution of this new function “transcends the phenotypic boundaries of a diverse and well studied species [E. coli].” The grounds for this statement are substantiated, as lack of growth on citrate under aerobic conditions has long been an established and defining characteristic of E. coli. Furthermore, utilization of citrate by the evolving population resulted in “an ecological transition from a single population to a two-member community,” since Cit– (non-citrate utilizing) E.coli remain and outcompete the Cit+ bugs for the low level of glucose in the media.
It certainly seems that criteria for evolutionary novelty have been met—but is it really novelty? Has a new species emerged from the LTEE?
When Is Something New Not Anything New?
The authors state, “A complete understanding of the evolution of a novel trait requires explanation of its ecological function, its physiological basis, the underlying mutations, and the history of the accumulated changes.” That seems to be exactly what they report in the initial and subsequent papers.5
So, does “novel” always mean something new? And in the case of this new trait, is it utterly new or a qualified new?
Well, it isn’t utterly new for E. coli to utilize citrate as a primary food source. E. coli utilizes citrate via a citrate fermentation cycle under anaerobic conditions just fine. Under anaerobic conditions, the bacteria produce a citrate/succinate antiporter that allows transport of citrate from the environment into the bacteria. Also, as mentioned before, citrate utilization by E. coli, although rare under aerobic conditions, has been observed before. In those instances, the acquisition of this novel function, like many acquisitions of new traits in microbes, is through the horizontal (from bug to bug) transfer of small circular pieces of DNA (known as plasmids) encoding new functional genes. They can grow aerobically on citrate because they produce a citrate transporter encoded by the plasmid DNA and regulated by plasmid promoters. The citrate transporter allows citrate uptake, and citrate is metabolized via the citric acid cycle.
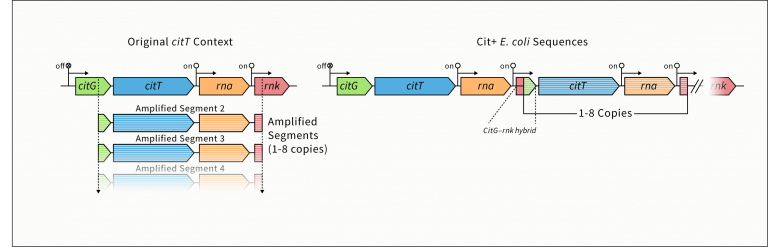
But that’s not what’s going on in the Cit+ E. coli in the LTEE. It’s not horizontal gene transfer. It’s not due to contamination of the cultures. The citrate transporter is not being co-opted from another system. It’s not exaptation. All Cit+ bacteria in the Ara-3 lineage harbor a ~3 kb tandem repeat of a portion of the endogenous citT operon, resulting in the introduction of additional citTgenes in the E. coli genome. (See figure below.) The critical (but not sole) changes hinge on the duplication of an endogenous promoter (rnk) that is active under aerobic conditions, the duplication of the citT gene, encoding a citrate transporter, and the juxtaposition of these elements in tandem repeats. The tandem copies of the citT gene are no longer under the regulatory control of the endogenous citG/citT promoter but are under the regulation of copies of an endogenous, downstream rnk promoter. The rnk-citT element allows the production of a citrate transporter under aerobic conditions, and this allows Cit+ E. coli to utilize the citrate in the growth media via mechanisms and metabolic pathways already present.
The Ara-3 Cit+ bacteria utilize citrate through the preexisting metabolic pathway (citric acid cycle). The ability to do so is facilitated by production of citrate/succinate antiporters from a preexisting gene sequence that is now expressed under aerobic conditions by simply switching the promoter that controls its expression. It is a novel trait, but the mechanisms underlying it are not innovations of new genes or new promoters. The Cit+ bacteria did not evolve a new transporter, a new promoter, or a new metabolic pathway.
Trying to Communicate Clearly—That’s Novel!
Perhaps now you can see how something novel isn’t really novel. The trait is novel, the genetic arrangement is novel, but the mechanisms, genes, promoters, and metabolic pathways are not novel. As it turns out, this highlights one more area where the ability to talk past one another, glossing over the word “novel,” in order to make one’s case is happening quite often in the evolution/antievolution exchange. No one is taking the time to try to understand each other or to clarify what’s really going on for the uninitiated.
When we dialogue with others who hold different perspectives, we must be able to listen first and then do the hard work of communicating thoughtfully and clearly. If we really value the truth and if we value and love the people we are conversing with, then we must take the time and make the effort to communicate well. It will be our motive and manner as much as our message that will impact others. For the sake of the gospel, let us make the effort to communicate truthfully and graciously, whether it’s about science or Scripture.
We can affirm that microbial evolution is taking place. Stepwise changes have accrued in a contingent fashion. Some E. coli have gained the ability to import citrate in the presence of oxygen. Under these experimental conditions, those bacteria that have evolved will thrive and continue to adapt. Biodiversity has increased, and an untapped ecological niche is now accessible. But we can also affirm that nothing analogous to macroevolution has truly happened here either. E. coli with or without a citrate transporter are still E. coli.
Zachary Blount, Christina Borland, and Richard Lenski, “Historical Contingency and the Evolution of a Key Innovation in an Experimental Population of Escherichiacoli,” Proceedings of the National Academy of Sciences, USA 105 (June 2008): 7899–906, doi:10.1073/pnas.0803151105.
Klaas Martinus Pos, Peter Dimroth, and Michael Bott, “The Escherichia coli Citrate Carrier CitT: A Member of a Novel Eubacterial Transporter Family Related to the 2-Oxoglutarate/Malate Translocator from Spinach Chloroplasts,” Journal of Bacteriology 180 (August 1998): 4160–65, https://jb.asm.org/content/180/16/4160.abstract.
Zachary Blount et al., “Genomic Analysis of a Key Innovation in an Experimental Escherichia coli Population,” Nature 489 (September 2012): 513–18, doi:10.1038/nature11514.